蛋白质分子是一种复杂的有机化合物,氨基酸是其组成的基本单位,是生命与细胞活动的重要组成部分。蛋白质种类成千上万,其复杂形态与精微结构,更是让科学界对其个体认识可谓捉襟见肘。由于科学界关于蛋白质分子的功能、模型结构介绍可谓汗牛充栋,这里作者同样不再赘述蛋白质分子的一般知识,只描绘蛋白质分子未被科学界所认识的以太波流一体运动与结构形态,及相关的拓展认识,同时也纠正其模型中的建立在经典错误原子模型上拓展出错误认识。
众多氨基酸分子,通过R基次生以太旋涡之间的耦合作用联结在一起成为一条长链分子,就是肽链。肽链周边空间也存在次生以太旋涡。由于以太旋涡运动会产生扭转力矩,当一条肽链独立于空间时,会让肽链产生扭曲结构与旋转运动,肽链上众多氨基酸的—CO—OH健由于流场相互作用的关系,产生范德华力而相吸并相互接近,让肽链最终产生团状结构,多条肽链耦合在一起,也是可以如此形成团状结构,这团状结构就是蛋白质分子。蛋白质分子周边空间也存在次生以太旋涡。
认识到以太存在,及振动力场与流场的作用形态,蛋白质分子的结构分类其实很简单。就如氨基酸分子以太旋涡其实是水分子以太旋涡的高阶变体,蛋白质分子,其实也只是氨基酸分子的高阶变体。
现代科学实验室发现生命体一共有22种氨基酸。众多氨基酸分子一共分三种以太波流一体形态,即酸性、中性、碱性,对应的是振动力场与流场的大小不同。应不同酸碱型或中性氨基酸的含量不同,在肽链上排位次序不同,肽链扭曲旋转后氨基酸在蛋白质空间的位置不同,组成的蛋白质分子有三种对应的形态,即酸型蛋白质分子,中性蛋白质分子,碱型蛋白质分子,及五种梯度分布下的蛋白质分子,即强酸型、弱酸型、中性、弱碱型、强碱型蛋白质分子。当然这里强酸弱酸、强碱弱碱只是蛋白质分子之间酸碱性的相对概念,不是无机物中强酸强碱这种PH值为1、2或PH值为11、12这样的强度下的说法,而是PH值范围为5-6即为强酸型,PH值范围为8-9即为强碱型,同样PH值范围为6-7是为弱酸型、PH值范围7-8是为弱碱型。这种数据是作者举例说明,具体强酸强碱或弱酸弱碱,则待科学界以后设定统一数据标准来参考。又应蛋白质分子中氨基酸数量的不同,分为大体积蛋白质分子,中体积蛋白质分子,小体积蛋白分子。这种酸碱性、体积大小相互组合,又会让相同酸碱性的蛋白质分子表达出不同的性状。这里作一一简单介绍其成因与功能。
1、酸型蛋白质分子
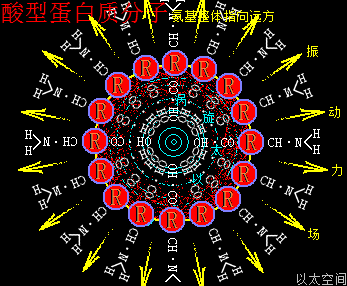
一类蛋白质分子,若其组成的氨基酸主体是由酸性氨基酸分子构成,那么众多酸性氨基酸分子的振动力场相互叠加强化,会让这类蛋白质分子对外作用表达为酸性,是为酸型蛋白质分子。在生命细胞的化学反应中,与无机化合物中的酸有相近的功能,主体起到分解与破坏作用。其以太波流一体状态下的力场对比为:
振动力场 > 流动力场
应酸性氨基酸分子所占比例不同,大比例强酸性氨基酸分子对应的是强酸型蛋白质分子,大比例弱酸性氨基酸分子对应的是弱酸型蛋白质分子。其它应氨基酸分子的强弱酸性不同与数量比例不同,有相应的强弱酸性梯度分布的酸型蛋白质分子。
强酸型蛋白质分子,其振动力场有最强的分解与破坏能力,对于大体积的强酸型蛋白质分子更是让这种能力达到极致,是催化酶的主体形式。会在后面章节专门介绍酶的催化作用的另类机制。弱酸型蛋白质分子可以活化细胞运动,是肌肉蛋白、血红蛋白等与运动相关组织蛋白的主体形式。

酸型蛋白质分子,其外部周边空间存在强振动力场,其内部空间除了存在氨基酸—R基团之间相互耦合吸引之外,其—CO—OH也大多集中在中心,相互之间产生范德华力吸引作用,进而强化这种耦合吸引作用,让蛋白质分子空间结构得以稳定,不会首先被自身的强振动力场分解与破坏。
酸型蛋白质分子的整体强振动力场向四周扩散,会对分子团上的个体氨基酸的—CH—NH2产生振动力场之间的排斥作用,使其指向远方,如此作用结果让蛋白质分子上大多氨基酸分子的—CH—NH2都朝外排列并指向远方,而—CO—OH则都朝内排列并指向中心。且—NH2在振动力场作用下除了指向远方之外,还让氮原子以太旋涡的黄道面涡流产生如伞面外凹偏向,让—NH2结构中的氮、氢原子核如一个树丫分叉般的排列。对于一个球形酸型蛋白质分子来说,如此—CH—NH2指向分布及—NH2分叉结构,其表面会看上去会有一个个双突点的结构形态,就如将许多小段树丫插满在一个大泥球上。
而—CO—OH朝内排列指向中心,其流动力场之间相互干涉排斥,在蛋白质内部空间形成一个以以太流场为主体作用方式的大场涡,即大场涡形成大以太旋涡,这以太旋涡偏向性与振动强度都很低,对内表达出很低的活跃性,犹如存在一个强流动力场。同时让蛋白质分子有聚合与收敛的趋势,在空间上蛋白质分子内部的各个氨基酸分子有更紧密的结合程度,如此导致酸型蛋白质分子的体积相对要小。
2、中性蛋白质分子
一类蛋白质分子,若其组成的氨基酸主体是由中性氨基酸分子构成,或虽由酸性氨基酸与碱性氨基酸构成,但酸性氨基酸与碱性氨基酸数量相当,相互的振动力场、流场能分别抵消,那么众多中性氨基酸分子为主体的蛋白质分子的振动力场与流动力场相互中和,会让这类蛋白质分子对外作用表达为中性。在生命细胞的化学反应中,这类蛋白质的功能与无机化合物中的水有相似的中和属性,主体起到构架与组织作用。其以太波流一体状态下的力场对比为:
振动力场 = 流动力场
中性蛋白质分子,由于振动力场与流动力场相当,在细胞中可被其它细胞器或组织结构所固定而不应自身活性原因脱离,故可以构架出稳定的细胞器的膜、管、腔等组织结构。不同体积大小的中性蛋白质分子对应不同大小的细胞器与组织结构。一般来说,大细胞器对应大体积中性蛋白质分子,小细胞器对应小体积中性蛋白质分子。由于中性并不是一个绝对概念,即不是测得PH=7,或振动力场刚好与流动力场完全抵消,而是有个偏差范围,偏差两端即为微酸与微碱,因此微酸型或微碱型蛋白质分子,也都可归于中性蛋白质分子一类。
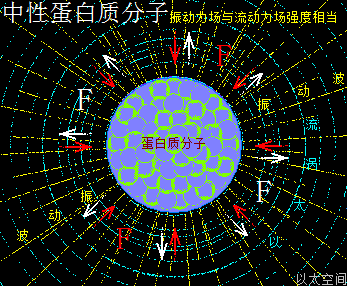
中性蛋白质分子,除了由中性氨基酸构成的纯中性蛋白质分子之外,应其构成的酸性氨基酸与碱性氨基酸的强弱不同,还有强酸—强碱型中性蛋白分子,弱酸—弱碱型中性蛋白质分子,共三种模式。这三种中性模式,就如无机分子中的纯水中性,氯化钠溶液中性,碳酸钙溶液中性类似,虽然各呈中性,但其各成分不同,其分子以太旋涡会对周边空间产生不同的波动形态,让以太处于不同强度与频率的氲氤状态,在宏观上会有不同的味道感官,同样,即便都是中性蛋白质分子,也是各自有不同的振动力场与流场的,需具体蛋白质分子具体考查。
特别是对于由强酸性氨基酸与强碱性氨基酸因数量相当、两种力场抵消所致的中性蛋白质分子,虽整体上表现为中性,但在这类蛋白质分子的局部区域仍保留强酸性与强碱性氨基酸的力场作用特征,会因外界其它分子的耦合作用,时而产生偏酸属性,时而产生偏碱属性的力场作用,并有对应的排斥或吸引现象,是膜运输蛋白质分子、免疫蛋白分子的主体成分。
在中性蛋白质分子内部空间,—CH—NH2与—CO—OH都有朝内排列并指向中心的,各自振动力场与流动力场之间相互干涉排斥并中和,在蛋白质内部空间形成多个涡轴不定,类似湍流形态分布的小场涡,同时让蛋白质分子空间有不稳定的趋势,会被外部其它分子以太旋涡的振动力场或流动力场所牵引而随时改变方位与轴自由度,及整体颗粒的定向位移。
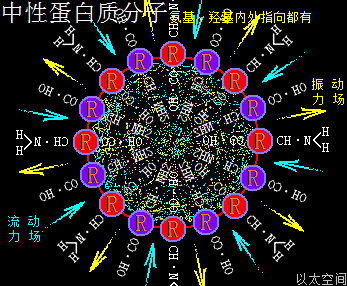
3、碱型蛋白质分子
一类蛋白质分子,若其组成的氨基酸主体是由碱性氨基酸分子构成,那么众多碱性氨基酸分子的流动力场相互叠加强化,会让这类蛋白质分子对外作用表达为碱性,这就是碱型蛋白质分子。在生命细胞的化学反应中,与无机化合物中的碱有相近的功能,主体起到合成与收敛作用。其以太波流一体状态下的力场对比为:
振动力场 < 流动力场
应碱性氨基酸分子所占比例不同,大比例强碱性氨基酸分子对应的是强碱型蛋白质分子,大比例弱碱性氨基酸分子对应的是弱碱型蛋白质分子。同样,其它应氨基酸分子的强弱碱性不同与数量比例不同,有相应的强弱碱性梯度分布的碱型蛋白质分子。
碱型蛋白质分子,由于其流动力场大于振动力场,可以有很强的吸附、收敛作用。对于大体积的强碱型蛋白质分子更是让这种能力达到极致,是染色体、染色质、核仁、叶绿体上的蛋白质的主体形式。弱碱型蛋白质分子可以吸附并固定其它营养分子,是与糖类、脂类、细胞核膜转运蛋白等与合成相关组织蛋白的主体形式。
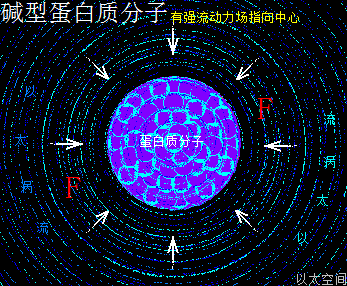
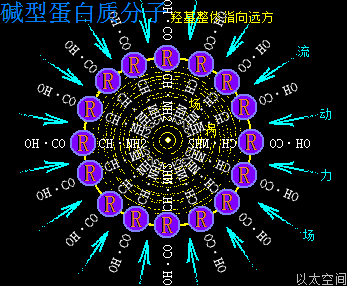
碱型蛋白质分子,其外部周边空间存在强流动力场,其内部空间除了存在氨基酸—R基团之间相互耦合吸引之外,其—CO—OH也大多分布在外围,相互之间产生范德华力吸引作用,进而强化这种耦合吸引作用,让蛋白质分子空间结构得以稳定,不会被内部的振动力场分解与破坏。
碱型蛋白质分子的整体强流动力场向内部收敛,会对分子团上的个体氨基酸的—CO—OH产生流动力场之间的排斥作用,将其推向远方,如此作用结果让蛋白质分子上大多氨基酸分子的—CO—OH都朝外排列并指向远方,而—CH—NH2则都朝内排列并指向中心。—CO—OH在流动力场作用下除了指向远方之外,还让氧原子以太旋涡的黄道面涡流产生如伞面外凸偏向。对于一个球形碱型蛋白质分子来说,如此—CO—OH指向分布,其表面会看上去会有一个个单突点的结构形态,就如将许多小段吸管插满在一个大泥球上。
而—CH—NH2朝内排列指向中心,其振动力场之间相互干涉排斥,在蛋白质内部空间形成一个以振动力场为作用方式的大场涡,即大场涡形成大以太旋涡,这以太旋涡偏向性与流转速度都很高,对内表达出很高的活跃性,犹如存在一个强振动力场。同时让蛋白质分子有分解与膨胀的趋势,在空间上蛋白质分子内部的各个氨基酸分子有更松散的结合程度,如此也导致碱型蛋白质分子的体积相对要大。
如此蛋白质分子按酸碱属性,可分五种形态:强酸型蛋白质分子,弱酸型蛋白质分子,中性蛋白质分子,弱碱型蛋白质分子,强碱型蛋白质分子。中性蛋白质分子又可分为强酸—强碱型中性蛋白质分子,弱酸—弱碱型中性蛋白质分子,纯中性蛋白质分子。这五种主体形态及三种细分形态是生命体蛋白质分子最重要的分类形态,由此人们不再被种类繁多、结构复杂、数量万千、大小不定的蛋白质分子所迷惑。会在后面章节继续论述以上几种形态的蛋白质分子的各种以太旋涡理论解析下的具体功能。
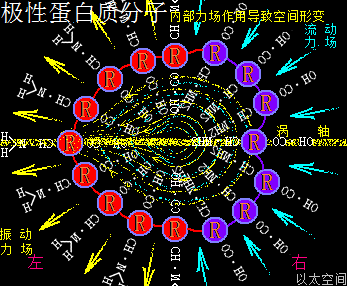
4、极性蛋白质分子
应包含各氨基酸的酸碱性数量不同,及各酸碱性氨基酸在蛋白质颗粒空间的疏密分布不同,导致蛋白质局部空间的振动力场与流动力场分布不均,从而让蛋白质分子次生以太旋涡表现出极性,这与极性分子以太旋涡的原理一样,只是构架空间要广大些,参与的微观以太旋涡要多些。在蛋白质分子空间里,由于氨基酸种类不一,力场不同,导致绝对均衡对称的氨基酸分子分布形态下的蛋白质分子结构是不存在的,于是可知所有蛋白质分子都是极性分子。其以太波流一体状态下的力场形态为:
振动力场、流动力场各自不对称
以一个球形蛋白质分子为例,设分子一半组成是强酸性氨基酸分子,分布在左半边,一半组成是强碱性氨基酸分子,分布在右半边,其振动力场与流动力场整体对外表现为中性,是为中性蛋白质分子。但在局部区域,即接近左半边顶点或右半边顶点的局部空间里,其振动力场与流动力场分别占主导地位。而接近左右半边交接处的局部空间里,其振动力场与流动力场又是相互中和。

如此结构的蛋白质分子,会应外界振动力场与流动力场的作用,而出现快速轴纠正反应。比如这蛋白质分子若出现在一个强振动力场环境空间里,蛋白质分子的振动力场与环境振动力场相互排斥,左半边会被推至背离环境振动源的方向,而右半边会被吸引至靠近环境振动源的方向,表达为轴纠正。反之这蛋白质分子若出现在一个强流动力场环境空间里,蛋白质分子的流动力场与环境流动力场相互排斥,右半边会被推至远离环境振动源的方向,而左半边会被吸引至接近环境振动源的方向,也表达为轴纠正。两种轴纠正都是极性蛋白质分子对环境变化作出的动作反应。即:
极性蛋白质分子会有轴纠正运动
若这个蛋白质分子处于氲氤状态下的以太空间里,由于左半边的振动力场以波的形式产生作用,会在左半边外围空间产生正以太压力,正以太压力会推动蛋白质分子向振动力场的反方向位移,也即右移动。同时,右半边的流动力场以流的形式产生作用,会在右半边外围空间产生负以太压力,负以太压力会牵引此蛋白质分子向流动力场的反方向位移,也即向右移动。两种力场都对此蛋白质分子产生向右的作用力,这蛋白质分子就会向右方向定向位移。自然,由于蛋白分子的振动力场及流动力场同以太空间相互作用的效果是很弱的,这种位移速度也是非常缓慢,但又是存在的。不同极性的蛋白质分子的运动形态也是千差万别的。这是极性蛋白质分子能够自主运动的力场作用根源,而非当下科学界对蛋白质分子的认识只是随机分布在空间的颗粒形态,可以任由环境变化摆布与安排。即:
极性蛋白质分子能够自主定向运动
极性蛋白质分子即能进行轴纠正运动,又能自主定向运动,因此对环境力场变化能随时随地产生空间位置变动与自身状态调整,这就是蛋白质分子的应激反应。
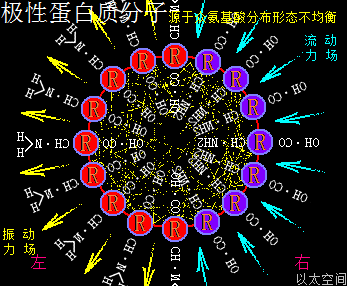
在这球形蛋白质分子内部空间,—CO—OH与—CH—NH2也是对应地处于分布疏密不均的状态,与外部的基键分布刚好相反,导致内部空间的振动力场与流场也是有单一指向性,是从—CH—NH2指向—CO—OH。而—CH—NH2的振动力场有向外扩张的趋势,—CO—O的流动力场有向内收敛的趋势,内部振动波从—CH—NH2指向—CO—OH并形成场涡,再牵引以太形成次生以太旋涡与涡管,涡轴是两力场的中心连接线。于是两个力场分别让—CO—OH聚集的区域向内收敛,又从涡管处向外扩张;让—CH—NH2聚集的区域向外膨胀,又从涡管处向内凹陷,形成—CH—NH2面钝,—CO—OH面尖的桃心形状的空间结构。这就是极性蛋白质分子空间形变的内在力场作用根源。其它更复杂的多氨基酸结合成的蛋白质分子,会也对应的更复杂的空间结构,但形变的作用原理都是一样的。
各酸碱性氨基酸应数量与种类不同,其不同排列组合形成的蛋白质分子可以有无穷多种,于是蛋白质分子的极性形态也是多种多样的。如有强酸—弱酸型·极性蛋白质分子,有强酸—中性型·极性蛋白质分子,有强酸—弱碱型·极性蛋白质分子,有强酸—强碱型·极性蛋白质分子;又有弱酸—中性型·极性蛋白质分子,弱碱—中性型·极性蛋白质分子,弱碱—强碱型·极性蛋白质分子,等等不一而足。又如有单涡轴极性蛋白质分子,有多涡轴极性蛋白质分子,等等。这些蛋白质分子的振动力场与流动力场在分子周边空间分布不均及强弱不同,并有各自特征,导致蛋白质分子对外界环境的不同振动力场与流动力场产生相应作用,表达为特定的功能。而所有功能,都是通过振动力场与流动力场产生作用的。